Karel Dorey Laboratory

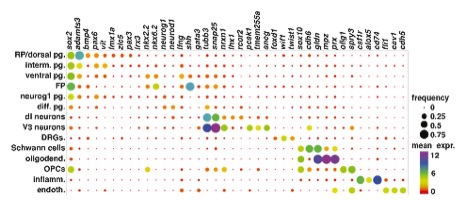
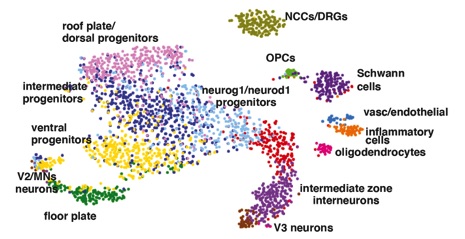
the cellular organisation of the Xenopus spinal cord is not well described, we first used the 10XGenomics platform to sequence 2908 cells from NF50 spinal cord. Twelve clusters were identified comprising the different cell types expected in the spinal cord: roof and floor plate (bmp4 and shh respectively), dorsal (lmx1a/zic5), intermediate (pax6, vit) and ventral (nkx2.2, nkx6.2) progenitors, neurons (tubb3, snap25), oligodendrocytes (mpz, prx) and oligodendrocyte progenitor cells (OPCs; sox2, olig1). Two clusters correspond to cells present in spinal ganglia: dorsal root ganglia (wif1 and twist1) and Schwann cells (sox10). Finally, we observed a small population of inflammatory (csf1r, alox5), vascular and endothelial (fli1, cav1) cells that may be spinal cord resident cells or a contamination from the dissection.
Organisation of the tadpole spinal cord
Regulation of axonal branching
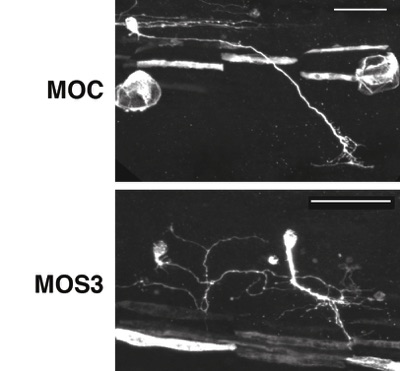
Spry3 expression in Xenopus embryos is restricted to the trigeminal nerve and both sensory and motor neurons in the spinal cord, and is dependent on BDNF-TrkB signalling. Loss-of-function experiments in Xenopus embryos show that Spry3 specifically represses the formation of new branches in motoneurons. In Xenopus spinal cord neurons in culture, loss of Spry3 expression causes an excess number of axonal filopodia in a BDNF signalling-dependant manner. Furthermore, in mouse cortical neurons, Spry3 over-expression reduces the number of branches induced by BDNF, indicating that Spry3 function might be conserved in mammals. Finally, Spry3 regulates specifically the Ca2+ pathway downstream of BDNF/TrkB signalling. Altogether, we show that Spry3 is a new regulator of axonal branching during embryonic development and a negative regulator of calcium signalling downstream BDNF-TrkB.